ISPP Task Force on Global Food Security
Meeting in Christchurch, New Zealand, 4-6 February 2003 within the 8th International Congress of Plant Pathology
Tuesday 4 February 2003, 09:00 - 12:00
Thursday 6 February, 09:00 - 12:00
Convention Centre, Meeting Room 2
SUMMARY RECORD
1. INTRODUCTION AND PLAN FOR THE MEETING
2. REVIEW OF FIVE CURRENT ACTIVITIES
ACTIVITY 1:
CHANGING PUBLIC POLICY AND OPINIONS ON GLOBAL FOOD SECURITY
1. Program
Symposium on Biosafety and Bioterrorism: Assessing the Threat to
Agriculture from Biological Agents
2. Student
Debate on the topic, Can we be prepared for deliberate release of biological
agents against agriculture?
3. Web
Forum on Biosafety
ACTIVITY 2:
ENHANCED PhD TRAINING FOR PLANT PATHOLOGY IN DEVELOPING COUNTRIES
ACTIVITY 3:
QUANTIFICATION OF THE ECONOMIC IMPACT OF SOME MAJOR DISEASES
ACTIVITY 4:
FARMER TRAINING IN SIMPLE DISEASE MANAGEMENT: PILOT PROJECT FOR CASSAVA IN GHANA
ACTIVITY 5: DEVELOPMENT OF THE ISPP WEBSITE
3. CONGRESS CHALLENGE
4. NEW INITIATIVES
5. ANY OTHER BUSINESS and NEXT MEETING
Present:
Chrys Akem - Department of Primary Industries, Queensland, Australia, chrys.akem@dpi.qld.gov.au
Charlie Delp - International Society for Plant Pathology, Tampa, Florida, USA, charliedelp@hotmail.com
Richard Falloon - New Zealand Institute for Crop & Food Research, Christchurch, New Zealand, falloonr@crop.cri.nz
Lodovica Gullino - University degli Studi di Torino, Italy, marialodovica.gullino@unito.it
Mike Jeger - Wye College, Kent, UK, m.jeger@ic.ac.uk
Hajime Kato - Professor Emeritus, Kobe, Japan, Hakatoh@aol.com
Chris Mundt - Oregon State University, USA, mundtc@bcc.orst.edu
Meena Muthumeenakshi - Horticulture Research International, Wellesbourne, UK, meena.muthu@hri.ac.uk
Serge Savary - ENSAR, Rennes, France, Serge.Savary@agrorennes.educagri.fr
Peter Scott - International Society for Plant Pathology, Oxford, UK, p.scott@cabi.org (Chairman)
Richard Strange - University College London, UK, r.strange@ucl.ac.uk
Tang WenHua - China Agricultural University, Beijing, China, tangwh@95777.com
Paul Teng - World Fish Center, Penang, Malaysia, p.teng@cgiar.org
Jacqui Wright - Secretariat of the Pacific Community, Suva, Fiji, jacquiw@spc.int
Apologies:
Jill Lenne - Milnthorpe, Cumbria, UK, jilllenne@compuserve.com
Emmanuel Moses - Crops Research Institute, Kumasi, Ghana, emoses@mail.com
Rebecca Nelson - Cornell University, New York, USA, rjn7@cornell.edu
C.Y. Shen - Food and Agriculture Organization of the United Nations (FAO), Bangkok, Thailand,
Chongyao.Shen@fao.org
Nollie Vera Cruz - International Rice Research Institute, Los Banos, Philippines, c.veracruz@cgiar.org
1. INTRODUCTION AND PLAN FOR THE MEETING
Peter Scott welcomed members of the Task Force and gave a brief background on the activities of the Committee, especially for the benefit of some new members that were participating for the first time. He then defined the agenda for the meeting. This was to continue the activities of the Task Force identified during the Workshop in Bangkok in 1999, to review submissions for the Congress Challenge, and to set a path for the future.
2. REVIEW OF FIVE CURRENT ACTIVITIES
ACTIVITY 1: CHANGING PUBLIC POLICY AND OPINIONS ON GLOBAL FOOD SECURITY
ISPP has been represented at a number of events at which it has aimed to effect changes in public policy and opinions on global food security.
- First Asian Conference on Plant Pathology, Beijing, August 2000
A paper was prepared by Dr Peter Scott for ISPP and Professor Tang WenHua (Beijing Agricultural University) on:
- Biotechnology in food security and environmental protection: The initiatives of the International Society for Plant Pathology and China's priorities.
It included sections on:
- Global food security: initiatives of the International Society for Plant Pathology
- Current status of biotechnology used in plant protection in China
Annexed was an ISPP Policy Statement:
Executive Committee of the International Society for Plant Pathology Supports Biotechnology
We, the Executive Committee of the International Society for Plant Pathology, meeting in Beijing, China, August 25-29 2000, believe and support the application of modern biotechnology as a tool to enhance agricultural productivity; to feed and improve the lives of the fast-growing world population and to address environmental degradation, hunger, and poverty. We also strongly advocate using sound science as the basis for regulatory and political decisions pertaining to biotechnology. We promote the careful, unbiased and science-based evaluation of technologies and products of modern biotechnology.
Modern biotechnology has tremendous potential and offers remarkable innovations to support efforts aimed at attaining food security. In particular, losses in quantity and quality of food caused by plant pathogens are known to be significant. Developing countries in general are expected to benefit more than developed countries from biotechnology in the areas of food and agriculture, health and medicine, environmental protection and trade and industry.
Products of modern biotechnology promise to reduce farmers' high input costs, increase their yields while helping conserve the ecosystem and enable crops to grow under normally unfavorable conditions. Biotechnology products can even provide greater benefits to consumers, who are the ultimate beneficiaries of technological innovations. For instance, modern biotechnology can be used as a tool to attain greater nutritional security through enhanced products qualities such as higher vitamin content, better protein quality and decreased post-harvest losses. Modern biotechnology can also produce healthier oils and develop vaccines to fight dreadful diseases like cholera and malaria.
While we recognize the importance of modern research, we must emphasize the new technology must be openly available (shared) as much as possible. We also recognize that no technology is without risk. However, we have great confidence in the safety of biotechnology conducted under appropriate protocols and in the products which undergo proper regulatory reviews before they are approved by governments for general use.
We therefore advocate and promote the safe and responsible applications of modern biotechnology in science and technology, agriculture and food, health and medicine, environment and trade and industry. Considering the tremendous potential of this technology, we urge policymakers to base their decisions on sound scientific evidence.
The Executive Committee of the International Society for Plant Pathology
This was adapted as a Statement "ASIAN PLANT PATHOLOGISTS IN SUPPORT OF BIOTECHNOLOGY" which was circulated at the Conference. It was signed by 110 of the 387 delegates, so 28% of the delegates (from 9 countries) expressed their support for ISPP's position.
The Congress included a Media Workshop on "Role of biotechnology for solving plant disease problems in sustainable agriculture". It was organized by Professor Tang WenHua, with the goal of influencing the policy debate in Asian countries by providing scientific information on key issues related to the role of modern technology in sustainable agriculture, with special emphasis on biotechnology. A debate was held between journalists from China, India and Malaysia and a panel of resource scientists: Dr. Peter Scott, CAB International, President of ISPP; Prof. Tian Po, Academician, China; Dr. Ed French, International Potato Center (CIP), Peru; Dr. Paul S. Teng, Monsanto Company - Asia Pacific; Dr. Tom Mew, International Rice Research Institute (IRRI ), Philippines. A number of programmes and articles were published as a result. Two sample titles are: China tightens controls on gene modified crops and More Asian nations to introduce transgenic crops
- Asian Agricultural Congress, Manila, April 2001
Professor Tang WenHua presented a paper by himself and Peter Scott entitled "Biotechnology in food security and environmental protection: The initiatives of the International Society for Plant Pathology and China's priorities". This was a development of the paper prepared for the First Asian Conference on Plant Pathology, and included a major section on the current status of biotechnology used in plant protection in China. The abstract read:
Biotechnology is a unique priority in the life sciences for the new millennium. The largest challenge that human beings face in 21st Century is food security. The first
"Green Revolution" in agriculture resulted in many high-yielding crops. Traditional knowledge and technology are still valuable to reduce poverty and food shortage. But with the increasing population new knowledge is needed and new technologies should be adopted. In this report, the current status of biotechnology used in plant protection in China is briefly introduced. The facts worldwide showed that biotechnology dramatically promoted development of agricultural sciences and technologies. The International Society for Plant Pathology actively encourages the development of biotechnology in agriculture for food security in this century. It can be proposed that biotechnology could be the most attractive approach to meeting the needs of the global population for food security. Some problems in the development of GM plants are discussed.
- World Food Summit: Five Years Later, Rome, June 2002
Dr Paul Teng was ISPP's representative. He circulated a submission paper prepared by ISPP Executive in consultation with ISPP Membership, reproduced here:
International Society for Plant Pathology (ISPP)
SUBMISSION
to the
World Food Summit: Five Years Later
10-13 June 2002, Rome
Introduction
The ISPP represents about 20,000 of the world's scientists who work in over 40 countries to control plant diseases which devastate crops and cause significant yield losses every year. The objective of the ISPP is to promote the worldwide development of plant pathology and the dissemination of knowledge about plant diseases and plant health management.
ISPP shares FAO's concerns about the underlying causes of world food insecurity and supports its attempts to facilitate change. In particular, ISPP notes with concern the need to reduce the losses caused by plant diseases in developing countries as part of the drive to increase crop productivity, and consequently the total supply of food at the local level.
"Global Food Security: The Role for Plant Pathology"
At its most recent International Congress of Plant Pathology (Edinburgh, 1998), ISPP held a public meeting on "Global Food Security: The Role for Plant Pathology", at which the enormity of the problem of food security, and its link with poverty, were noted. This meeting supported the determination of the World Food Summit in Rome,1996, to halve the number of hungry people by 2015.
The impact of pests, diseases and weeds on food supply was highlighted, noting estimates that they reduce production by at least one-third, and that diseases alone reduce production by more than 10%. The losses due to various biotic stresses are greater in the developing countries. If the post-harvest losses are also included the situation becomes even more alarming. The Sanitary and Phytosanitary (SPS) Agreement of the World Trade Organization, aimed at developing measures that would ensure protection of human, animal and plant life or health, and transparency in global agricultural trade, adds another dimension to the urgent need to address plant disease problems. There are also serious concerns related to the impact of climate change and indiscriminate use of pesticides on pest and disease problems.
Task Force on Global Food Security
Options for managing crop diseases to improve food security were addressed at the 1998 Congress by distinguished scientists, including Nobel Peace Laureate Norman Borlaug.
ISPP then set up a Task Force to focus its activities, and concentrate on delivering some tangible results of demonstrable value. The Task Force represented plant pathologists from many different regions and disciplines.
Six areas were identified in which ISPP might reasonably effect useful change:
1 Policy; 2 Education; 3 R & D; 4 Farmer-practice; 5 Public opinion;
6 Information.
These topics were shared with representatives of National Programmes in Asia, FAO and other bodies represented by CROPLIFE Asia and GFAR (Global Forum for Agricultural Research). Specific actions have been proposed:
1. Establish an ISPP Policy Group
With a programme to inform policy makers of the importance of plant diseases in food insecurity
2. Study PhD training for plant pathologists in developing countries, with a view to enhancement
To assist decision making by donors, national agricultural research services, and universities in developing and developed countries
3. Quantify economic impact of some major diseases
To provide a record of major examples of the consequences of diseases for food security, pointing to the need for support for continuing research
4. Develop a pilot project for farmer training in simple disease management
Focused on basic knowledge of cassava diseases in Ghana, to help in diagnosis and management, as a starting point for further knowledge transfer
5. Develop the ISPP Website as a means of improved communication
To provide information about ISPP, plant pathology, and the work of the Task Force
All the proposed Activities are considered to be: 1 Appropriate to the status of ISPP;
2 Achievable within a time frame of 3 years; 3 Capable of being resourced;
4 Likely to deliver tangible results that will effect useful change. All are in progress. Developments are reported on the ISPP Website <http://www.isppweb.org>
The above activities are presented to the World Food Summit: Five Years Later FOR DISCUSSION AND ENDORSEMENT . . .
. . . as an example of some specific contributions a professional society may make to the challenge of global food security. With such endorsement, they can be reassessed and refined at the next International Congress of Plant Pathology (Christchurch, New Zealand, February 2003).
In addition, ISPP RECOMMENDS . . .
. . . that the following actions may be taken by National Agricultural Systems (NARS), particularly in developing countries, for reducing poverty, hunger and malnutrition through sustainable increase in agricultural productivity:
a) Provide a significant increase in long-term funding for agricultural research
b) Provide agricultural scientists with modern communications in order to access the latest scientific information relevant to their field of research
c) Strengthen undergraduate and graduate teaching in agriculture, including plant protection, so that a science-based research community can contribute to the generation of the knowledge required to facilitate food security
d) Provide a modern infrastructure of technological empowerment of farmers in order to enhance food security
and
e) Increase emphasis on plant protection research, so that low-cost technologies can be made available to farmers, to reduce losses caused by pests and pathogens.
Peter Scott
President, International Society for Plant Pathology
c/o CAB International, Wallingford OX10 8DE, UK. Tel +44 1491 832111
p.scott@cabi.org
Paul Teng reported that the ISPP paper was submitted for distribution but that it was difficult to assess what impact it had or which of its contents were incorporated into the Summit's statements and proceedings. This was because there were literally hundreds of submissions and only official government delegates were accorded the right of making comment on official Summit documents.
The role of science and technology was generally endorsed by the Summit, although there was caution on the use of biotechnology. The final text issued at the Summit included this statement:
The introduction of tried and tested new technologies including biotechnology should be accomplished in a safe manner and adapted to local conditions to help improve agricultural productivity in developing countries. We are committed to study, share and facilitate the responsible use of biotechnology in addressing development needs.
• ISPP and Entomological Society of America: Events on BIOSAFETY, Fort Lauderdale, November 2002
Dr Ernest Delfosse for the Entomological Society of America and Dr Peter Scott for ISPP organized three events focusing on Biosafety, stimulated by the terrorist events of September 11, 2001:
1. A Program Symposium on Biosafety and Bioterrorism: Assessing the Threat to Agriculture from Biological Agents
2. A Student Debate on the topic, Can we be prepared for deliberate release of biological agents against agriculture?
3. A Web Forum on Biosafety.
The Program Symposium included presentations on: Research gaps in systematics of potential arthropod biological agents by Michael E. Schauff, USDA-ARS; Research gaps in systematics of potential pathogen biological agents by Amy Y. Rossman, USDA-ARS; What are the other gaps in the American safety net? by Charles Schwalbe, USDA-APHIS; Deliberate or accidental? A proactive or reactive response? What do we need to do to prepare for new introductions of biological agents? by Ian McDonell, North American Plant Protection Organization; Risk analysis of biological agents that could be used to disrupt food and fibre production by Ernest S. Delfosse, USDA-ARS.
The Student Debate, which immediately followed the Program Symposium, was a closely fought contest between two teams of four students drawn from seven US universities, four from plant pathology departments and four from entomology departments. The outcome, determined by the audience in a secret ballot, was a narrow preference for the presentation by the team asserting that "We can be prepared for deliberate release of biological agents against agriculture".
The Web Forum, which ran from 1 to 15 November 2002, attracted 85 postings on a wide range of topics. Contributions have been archived and can be viewed by clicking on the link www.entsoc.org/biosafety. Principal topics included: The quarantine inspector's dilemma; PRA: an art or a science?; Are malicious introductions that big a threat?; Is it riskier to release GM plants than non-GM plants?; Is it really a food problem or an economic issue?; Plant health lacks the terror factor; APS is greatly concerned about biosecurity; Objectives of terrorists. The Web Forum was technically managed by Dr Terry Stewart of Massey University, Palmerston North.
The three events were complementary and produced significant interest in a topical issue. Combinations of this sort may be appropriate in the future to address other topics in food security.
• Plant Disease: A Threat to Global Food Security, Chapter in Annual Review of Phytopathology, 2005
This review is reproduced by kind permission of Annual Reviews. It can also be accessed at the
Annual Review of Phytopathology website and, in hard copy, in Annual Review of Phytopathology Volume 53, pages 83-116 (2005), obtainable from Annual
Reviews, 4139 Camino Way, PO Box 10139, Palo Alto, California 94303-0139, USA.
www.annualreviews.org
Annual Review of Phytopathology Vol. 43: 83-116 (Volume publication date September 2005) (doi:10.1146/annurev.phyto.43.113004.133839)
PLANT DISEASE: A Threat to Global Food Security
Richard N. Strange
Department of Biology, University College London, London WC1E 6BT, United Kingdom; email: r.strange@ucl.ac.uk
Peter R. Scott
CAB International, Wallingford OX10 8DE, United Kingdom; email:
p.scott@cabi.org
ABSTRACT
A vast number of plant pathogens from viroids of a few hundred nucleotides to higher plants cause diseases in our crops. Their effects range from mild symptoms to catastrophes in which large areas planted to food crops are destroyed. Catastrophic plant disease exacerbates the current deficit of food supply in which at least 800 million people are inadequately fed. Plant pathogens are difficult to control because their populations are variable in time, space, and genotype. Most insidiously, they evolve, often overcoming the resistance that may have been the hard-won achievement of the plant breeder. In order to combat the losses they cause, it is necessary to define the problem and seek remedies. At the biological level, the requirements are for the speedy and accurate identification of the causal organism, accurate estimates of the severity of disease and its effect on yield, and identification of its virulence mechanisms. Disease may then be minimized by the reduction of the pathogen's inoculum, inhibition of its virulence mechanisms, and promotion of genetic diversity in the crop. Conventional plant breeding for resistance has an important role to play that can now be facilitated by marker-assisted selection. There is also a role for transgenic modification with genes that confer resistance. At the political level, there is a need to acknowledge that plant diseases threaten our food supplies and to devote adequate resources to their control.
INTRODUCTION
More than 800 million people do not have adequate food; 1.3 billion live on less than $1 a day and at least 10% of global food production is lost to plant disease (20, 34, 63). Plant pathologists cannot ignore the juxtaposition of these figures for food shortage and the damage to food production caused by plant pathogens.
Fourteen crop plants provide the bulk of food for human consumption (Figure 1). In addition, there are numerous plants that are less intensively grown yet fulfil important nutritional requirements in restricted areas of the globe. All are subject to disease both in the field and post-harvest, the major groups of pathogens being viruses, bacteria, Oomycetes, fungi, nematodes, and parasitic plants. These biotic constraints can, at times, seriously compromise food security. For example, potato blight, caused by Phytophthora infestans, struck Europe like "a bolt from the blue" in the 1840s. In Ireland, about a million people died of starvation and more than a million attempted to emigrate (80, 117). The reasons for this calamity were the arrival in Europe of a virulent strain of the pathogen, the high dependence of much of the Irish population on potato for sustenance, the lack of resistance in the plant to the pathogen, and weather conditions favorable to epidemic development. There have been other disasters caused by plant diseases such as the Great Bengal Famine of 1943 (98) and the southern corn leaf blight epidemic of 1970–1971 in the USA (130), to name but two. In the former, an estimated 2 million people died owing to the high dependence of most of the population on a single crop, rice, which was attacked by the fungus Cochliobolus miyabeanus. In the USA, by contrast, although in some areas the corn (maize) crop was completely destroyed by another fungus from the same genus, Cochliobolus heterostrophus, alternative sources of nutrition were plentiful so no one died, although the effect on the agricultural economy was severe.
|
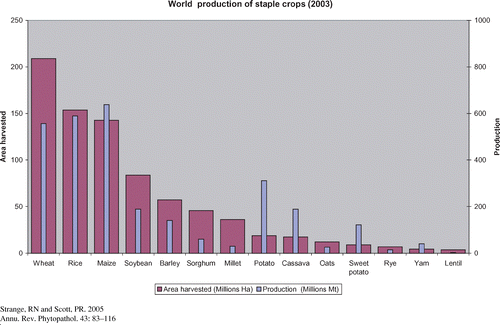
|
|
Figure 1 World production of staplea
crops.
|
The first two of these painful examples demonstrate with brutal clarity that in areas of the world where a large proportion of the population is dependent on a single crop or a few crops, they are at risk should that crop fail owing to one or more devastating diseases. At the present time, the threat is particularly great in developing countries, where populations are growing fastest, poverty is endemic, the population depends on locally produced staples, and the infrastructure of extension and R & D is often poorly resourced.
Globalization of agriculture has meant that crop plants, often with a narrow genetic base, are now grown far from their centers of origin and therefore far from the pathogens that have coevolved with them. As a result, they are unlikely to have evolved resistance to new strains of the pathogen that may have subsequently arisen in the center of origin. Should these new strains migrate to the new area of cultivation, as may have happened with P. infestans in the 1840s, the crop would be susceptible. Moreover, crops introduced to a new area may be poorly equipped to resist pathogenic organisms that may be resident there.
The problem of plant disease, particularly in developing countries, is exacerbated by the paucity of resources devoted to their study. In part, this may be a result of poor governance but it also arises from the difficulty of quantifying plant disease and relating this to the failure of crops to reach achievable yields. The inability to supply such hard data to administrators and politicians has meant that plant pathology, in relation to its importance, continues to be grossly underfunded. We hope that this review will help plant pathologists, particularly those in developing countries, to focus on the acquisition of such data. Moreover, once armed with hard statistics, it is hoped they will be more successful in securing adequate and appropriate funding from governments and other agencies in the public and private sectors in order to tackle effectively the plant diseases that destroy their most important crops.
Other difficulties faced by plant pathologists are the reliable identification of the causal agents of a disease to levels—species, formae speciales, pathovars, biovars and races—that are appropriate to their properties as plant pathogens and therefore relevant to their control; identification of sources of inoculum; the wide host range of some pathogens; lack of availability of sources of durable genetic resistance; and the slow acceptance in some cultures of the value of transgenic traits in plant improvement. Tropical climates, prevalent in most developing countries, cause particular difficulties in that they allow continuous cropping with the consequent build-up of inoculum. In contrast, the winters of temperate climates reduce inoculum levels owing to the reduced availability of hosts and inclement weather.
Solutions to the problems of plant disease come from many quarters. Exclusion of the pathogen through plant quarantine is the first line of defense and deserves more resources in relation to food security in developing countries. Other solutions include the exclusion, elimination or reduction of the pathogen's inoculum and development, for example by good cultural practices; intercropping and rotation; the judicious use of pesticides; exploiting the gene pool of the plant and its relatives in breeding programs (often the preferred approach where it is practical); understanding and combating virulence mechanisms of the pathogen; biological control (often difficult for plant pathogens); post-harvest protection; and improving plant performance through biotechnology. Current cultural reservations in public opinion about biotechnology should not inhibit its imaginative application to the challenging demands of food security, especially in developing countries. Maintenance of genetic diversity in crop plants has an overriding importance.
In this review, some examples of organisms that pose the greatest hazard to the successful cultivation of the world's most important edible crop plants are considered. Areas covered are pathogen identification, disease severity, epidemiology, and the evolution of virulence. The discussion then moves to options for control. In the first instance, these center on the exclusion and elimination of pathogens and, where these have failed, crop husbandry practices that limit the impact of disease. An important factor in control is the availability of resistant plants with favorable agronomic characteristics, and therefore the means by which they may be generated will be treated, including marker-assisted selection (MAS) and exploitation of knowledge of the biochemistry and molecular biology of plant-pathogen interaction. Finally, consideration is given to the feasibility and desirability of using biotechnology to ensure that sufficient food of high quality is available to all in the twenty-first century. Because this is a review of plant disease, we have given little attention to biologically caused post-harvest losses—though these have a major additional effect on global food security, and the problems they cause deserve more research and development than they receive. At the political level, the International Society for Plant Pathology has established a Task Force on Global Food Security. The challenges that it addresses attract less attention than those of global terrorism in the current climate of world opinion, but they present threats of a different degree, which need to be addressed through science-based co-operation on a global scale.
WHAT ARE THE THREATS?
The Crop Plants that are the Principal Sources of Human Nutrition and their Most Important Pathogens
These are given in Table 1. Although this is not the place to give a detailed account as to how each pathogen threatens food security, some examples of diseases caused by the major types of pathogens have been chosen to demonstrate the vulnerability of some crop plants that are of global importance or of critical importance to communities living in more restricted areas.
|

|
TABLE 1 The world's staplea crops and their principal diseases
|
|
Virus Diseases
Many of the more than 700 known plant viruses cause devastating diseases and often have wide host ranges. Barley yellow dwarf viruses (BYDV), for example, are distributed worldwide and infect over 150 species of the Poaceae, including most of the staple cereals—wheat, barley, oats, rye, rice, and maize (Table 1). A number of studies have demonstrated the debilitating effect these viruses have on cereals. For example, grain yield of maize infected with the PAV serotype of the virus was 15%–20% less than controls because infected plants have fewer kernels per ear (12), whereas in barley, infected with the same serotype, reductions of up to 38% were recorded (29).
Cassava is a very important food security crop for smallholder farmers, particularly in low-income, food-deficit countries, owing to its reputation for reliability (8). World production from 2000 to 2003 averaged 185 million tons/year with just over 100 million tons harvested in Sub-Saharan Africa (35). However, the plant is subject to diseases caused by mosaic viruses. Originally there were thought to be two strains, African cassava mosaic virus (ACMV) and East African cassava mosaic virus (EACMV), but in the late 1980s an unusually severe form of the disease was reported from Uganda. This was caused by double infection with a recombinant virus derived from ACMV and EACMV, termed UgV, and one of the parental strains, ACMV (139). In 1999, Legg reported that the area affected by the severe form of the disease had expanded to cover virtually all Uganda and large parts of Kenya, Tanzania, Sudan, and the Democratic Republic of Congo (85). Also, such was the sensitivity of local cultivars and the consequent losses that many farmers had abandoned cassava cultivation, consequently destabilizing food security in East Africa (85). More recently, the disease has spread to Rwanda (86), Burundi (14), and Gabon (87).
Diseases of cassava caused by mosaic viruses are not known in South America, the center of origin of the crop (36). The African experience is therefore a good illustration of the hazard of exporting a crop to another country where it may fall victim to a pathogen acquired from the local flora.
Bacterial Diseases
Species of bacteria belonging to several genera are devastating plant pathogens. For example, at least 350 different plant diseases are caused by species of Xanthomonas (88). One of these is bacterial blight caused by Xanthomonas oryzae pv. oryzae (Table 1). It is a major disease of rice in tropical Asia and all varieties of basmati rice are highly susceptible. Unfortunately, the pathogen is present in the entire basmati rice-growing area in India and, in consequence, production is severely constrained (70).
Recently, a severe disease of banana has been reported from Uganda (129). The symptoms were rapid yellowing and wilting of younger leaves, discoloration of the internal vascular vessels, and occasional dieback and internal rotting of banana fruits. These symptoms are very similar to moko disease caused by Ralstonia solanacearum, but this pathogen had not previously been recorded on banana in Africa. A bacterium was isolated from diseased bananas and proved to be the causal organism by the application of Koch's Postulates. Identification was made by comparing DNA fingerprints of the bacterium obtained from rep-PCR, using ERIC and BOX primers (90), and by showing that these matched fingerprints obtained from authentic cultures of X. campestris pv. musacearum, originally isolated from Ensete and Musa in Ethiopia (129).
Ralstonia (Pseudomonas) solanacearum, a plant pathogen with a worldwide distribution, causes diseases in more than 200 host species belonging to 50 families including potato (112) (Table 1). It exists as five races with different hosts and geographic distributions. Race 1 has a very wide host range and is endemic in the southern United States. Bananas are the principal host of race 2, in which, as noted above, it causes moko disease. This race is present in the Caribbean, Brazil, and the Philippines. Race 3 is distributed worldwide with the exception of the United States and Canada and infects many members of the Solanaceae such as potatoes, tomatoes, eggplant, and peppers, as well as geraniums. Ginger is affected by race 4 in much of Asia and mulberries in China by race 5. The organism is further differentiated into biovars on the basis of biochemical tests, e.g., race 3 biovar 2, which is present in Europe, Asia, South and Central America, and Australia (22; http://www.aphis.usda.gov/ppq/ep/ralstonia/background.html). Recently, this organism was detected in the United States in some greenhouses that had imported geranium plants from Kenya. The identification was confirmed in February 2003 and the organism eradicated. However, it was detected again in late December 2003, and the facility in Guatemala where the infected geraniums originated has stopped shipping to the United States (
http://www.aphis.usda.gov/ppq/ep/ralstonia/background.html ).
Fungal Diseases
There are at least five reasons why fungi may cause catastrophic plant disease:
|
1.
|
Many sporulate prolifically, the spores providing copious inoculum, which may infect further plants.
|
|
2.
|
Their latent period, i.e., the time between infection and the production of further infectious propagules, usually spores, may be only a few days.
|
|
3.
|
The spores, if they are wettable, may be spread as high-density inoculum in surface water or in droplets by rain-splash. Alternatively, nonwettable spores may be carried long distances by the wind.
|
|
4.
|
They may produce compounds that are phytotoxic and/or a battery of enzymes that destroy the plant's structure.
|
|
5.
|
Pathogens may draw nutrients away from the economically valuable part of the plant by the production or induction of growth regulators, such as cytokinins, and consequently depress yields (117).
|
Colletotrichum gloeosporioides (anamorph) or Glomerella cingulata (teleomorph) are names given to an organism that causes diseases, often referred to as anthracnoses, in many tropical and subtropical crops. Considerable variation occurs in culture and host range, with some strains able to attack many host species whereas others, such as those infecting mango, are confined to a single species (52). This variation has cast doubt on whether C. gloeosporioides can really be considered as a single species, and it has been suggested that it is a species complex (119). Here, molecular techniques are invaluable in defining strains. For example, molecular approaches demonstrated that C. gloeosporioides infecting Stylosanthes in Australia consisted of two clonal populations that do not combine readily in the field and that probably resulted from two separate introductions into the country (94). Similarly, isolates from yam (Dioscorea alata), a weed (Mikania micrantha), kava (Piper methysticum), and pigeonpea (Cajanus cajan) obtained from islands in the South Pacific were identical when compared by Amplified Fragment Length Polymorphism (AFLP), suggesting that they were clonal. Also, isolates from cassava and coffee obtained from Hainan Island, China, produced identical AFLP profiles, but these were different from the South Pacific profile and the profile obtained from a strain isolated from infected cassava in Ghana (2).
The wide host range of some strains of C. gloeosporioides presents a considerable threat to crops growing in countries where there is no fallow period corresponding to the winter of temperate climates. Even if time is allowed to elapse between the plantings of susceptible crops, the fungus may survive on weed species. Moreover, care has to be taken, if intercropping with other species is practiced, that these are not also susceptible to the prevalent strain of C. gloeosporioides.
Rice is second only to maize in global production (Figure 1) and is more important since it is a staple for about half the world's population, among whom are many of the poorest. It is attacked by the Ascomycete fungus Pyricularia oryzae [teleomorph Magnaporthe grisea], causing rice blast, resulting in losses of 10%–30% of the crop every year (121). To give one example, more than 700 ha of rice of diverse genotypes with varying levels of resistance in Bhutan were affected in 1995, resulting in losses of 1090 tonnes (126). Other cereals are also affected by P. oryzae or the similar species P. setariae; these include finger millet, Eleusine coracana, which, when attacked before grain formation, can suffer complete loss of yield (30) (Table 1). Such an attack has serious consequences in India and southern and east African countries where finger millet is an important food security crop. A species of Pyricularia has also been reported as an increasingly important problem on wheat, and serious outbreaks have occurred in Paraná, Brazil (61).
Several fungi that infect staple foods before and/or after harvest produce powerful mycotoxins. For example, the fumonisin toxins were discovered as a result of an investigation into the high level of esophogeal cancer in the Transkei region of South Africa. They were isolated from cultures of Gibberella fujikuroi (anamorph Fusarium moniliforme) grown on maize, and the most active compound was designated fumonisin B1 (FB1) (42). FB1 is a sphinganine analogue that, in both plant and animal cells, competitively inhibits sphingolipid biosynthesis, causing sphingoid bases to accumulate (95). High levels of virulence for maize were always associated with strains of the fungus that produced fumonisins (27).
Oomycete Diseases
Although the Oomycetes share the first four characteristics of fungi enumerated above that make them such dangerous pathogens of plants, they differ from them in several important features. These include the absence of chitin from their cell walls, the negligible sterol content of their membranes, their predominantly diploid karyotype, and their biflagellate zoospores. Nucleic acid and protein sequence data have now shown that they are members of the Kingdom Stramenopila and are more closely related to the golden brown algae than to true fungi (5). An important consequence of their independent evolution of the plant parasitic lifestyle from that of fungi is that some plant defenses such as the production of saponins and chitinases, which specifically target the sterols of fungal membranes and chitin, respectively, are ineffective against the Oomycetes.
The genus Phytophthora (meaning plant destroyer) is an Oomycete, and P. infestans, mentioned in the Introduction, is perhaps its most notorious species, costing annually on a global basis in excess of $5 billion in terms of losses of the potato crop and control measures (15). Pythium is another genus that can cause serious disease problems. For example, P. aphanidermatum and P. myriotylum cause severe root rot in bell pepper with 42% and 62% plant mortality, respectively (19). Similarly, members of the downy mildews such as Peronosclerospora, Peronospora, Pseudoperonospora, Plasmopara, Sclerophthora, and Sclerospora regularly cause severe diseases on a variety of crops that include maize, sorghum, pearl millet, onion, soybean, cucurbits, and grapes (125) (Table 1).
Nematode Diseases
Of the 17 orders of nematodes, only two, the Tylenchida and the Dorylaimida, contain members that are plant parasites (135). However, those that are parasitic may cause serious crop losses. For example, Ditylenchus dipsaci attacks over 450 different plant species, including weeds, and is one of the most devastating nematode species (45). Similarly, species of Meloidogyne such as M. hapla also attack many important crop plants, including groundnut, potato, carrot, sugarbeet, strawberry, and onion, and on occasions, may cause total crop loss (32, 91). Other species such as the soybean cyst nematode (Heterodera glycines) are more specific. Often its direct effects on crop losses are not noticed in the field but yields may be severely depressed. For example, losses in the southeastern United States were estimated at $88.4 million in 1990 (113). Nematodes that are members of the Dorylaimida may also act as vectors for viruses. For example, Tobacco rattle virus, which causes considerable economic damage to vegetable and ornamental flower-bulb crops, is efficiently transmitted by Paratrichodorus anemones (72).
Diseases Caused by Parasitic Plants
There are over 3000 species of parasitic plants among which Striga and Orobanche are, perhaps, the most notorious (116). Species of Striga infect more than two thirds of the 73 million ha of cereals and legumes grown on the African continent, affecting the lives of over 100 million people in 25 countries. Losses may be total, and infestation of some areas, owing to the enormous seed production by the pathogen, may be so great that the cultivation of some crops has to be abandoned (31). Broomrapes (Orobanche species) are parasitic weeds that are widespread in many countries of Central Asia and the Mediterranean region (65). They infect the roots of dicotyledonous crops and, because they do not contain chlorophyll, derive their entire nutrient requirements from their hosts. An aerial shoot is produced that may support several hundred flowers, each containing around 500 seeds, but by this time, the health and productivity of the host plant has been severely compromised. Like Striga, these pathogens may remain dormant in the soil for >10 years but still cause serious economic losses if susceptible crops are planted (24).
HOW SERIOUS ARE THE THREATS?
To evaluate the seriousness of a plant disease it is essential to identify the pathogen, determine the effect it is causing, identify the sources of inoculum, and gain some knowledge of the means by which it is spread. Although these stipulations appear elementary, they are frequently ignored because of lack of personnel and facilities for such work. In consequence, the disease develops and spreads unchecked. Such problems are most severe in developing countries, sometimes reflecting poor governance but also the lack of availability of resources to identify and quantify disease outbreaks and to relate them to the failure of crops to reach achievable yields. In the absence of a clear presentation of this relationship, plant pathology is likely to remain severely underfunded in relation to its importance for food security—a problem in both industrialized and developing countries.
Knowing the Enemy—Pathogen Identification
The correct identification of plant pathogens can hardly be overemphasized since it is fundamental to control. Initially, pathogens are often identified by the symptoms they cause. Although such identifications can be reliable when staff are experienced, they can frequently be wrong when such experience is not available. For example, it is difficult to avoid the conclusion that some formae speciales of Fusarium oxysporum are diagnosed purely on the basis of the vascular discoloration and wilts they cause, coupled with the name of the plant from which they were isolated. Indeed, recently a highly toxigenic isolate of a Fusarium obtained from chickpea was demonstrated by its morphology and rDNA sequences to be F. acutatum rather than F. oxysporum f. sp. ciceri (S. Gopalakrishnan & R.N. Strange, submitted).
Fortunately, a number of aids are available to plant pathologists with minimal facilities. At the field level there is the CABI Crop Protection Compendium (24), which provides a simple basis for preliminary identification through narrowing down the possibilities in a relational database of potential causes. The dimensions in the relational database allow the user to specify (a) the crop (say rice: of nearly 4000 plant pathogens listed in the Compendium, some 114 are recorded as affecting rice as a major host); (b) the country (say India: this reduces the number of possibilities to some 72); (c) the part of the plant affected (say the leaves: this reduces the number of possibilities to some 43); and (d) the symptom (say yellowed or dead leaves: this reduces the number of possibilities to some 8). A conclusion is reached in a few seconds. Each item in the final list is accompanied by a thumbnail illustration, and then leads to a full description of the condition, its cause, how damaging it is, and what can be done about it, together with illustrations and distribution maps.
At the laboratory level, many techniques are now available for definitive identification of plant pathogens, ranging from those that are simple and do not require expensive apparatus to the complex, which require costly equipment (reviewed in 107). Enzyme Linked Immunosorbent Assay (ELISA) is the preferred technique of diagnostic laboratories for bacteria and viruses because of its high throughput capacity (50, 105, 109), but PCR (polymerase chain reaction) techniques are rapidly becoming established. In the Laboratory Guide for Identification of Plant Pathogenic Bacteria (109) many primers for important plant pathogenic bacteria are listed. Real-time PCR is increasingly being used but the cost is prohibitive for many countries where it could be of most benefit. However, some lower-cost portable thermocyclers have been developed that can give real-time results in the field. For example, both watermelon fruit blotch, caused by Acidovorax avenae subsp. citrulli, and Pierce's disease of grape, caused by Xylella fastidiosa, were diagnosed in 1 h or less on-site (108, 110, 111).
PhytID (99) offers a molecular key to 50 species of Phytophthora, for which accurate identification is crucial to plant health legislation and plant quarantine issues. The system is based on the Internal Transcribed Spacer (ITS) regions of ribosomal RNA. Digest patterns are entered into an analysis package or simply compared with samples for different species. If a match can be found, unknown isolates may be identified in a matter of hours rather than the several days required for traditional morphological analysis.
If a known organism has been reliably identified as the cause of disease, then referral to the literature will often give invaluable help in evaluating the risk it poses. The Crop Protection Compendium (24) includes an Economic Impact module, and its descriptions of each of the major diseases include an expert assessment of the damage they do.
Rapid detection is of the essence and may be the key to success in containment and control (107). However, any doubt about the identity of the causal agent should be removed by the application of Koch's postulates, suitably modified in the case of obligate pathogens.
Measurement of Disease and Its Effect on Yield
The measurement of disease continues to be an area fraught with difficulty. In the first instance, plants are irregularly shaped three-dimensional objects. Second, the symptoms caused may range from a few isolated lesions to wilting and collapse of the whole plant. Third, there is usually no simple relationship between measures of symptoms and the failure to reach achievable yields (117). Although image analysis may be of help in obtaining an objective measure in some instances, e.g., with detached leaves bearing discrete lesions, it is generally of little help in evaluating disease in the field, particularly when this involves hundreds of lines in a trial. Here the time-honored practice of scoring on an ordinal scale, often of 1–9, is still the most common approach. However, little credence can be placed in such scores when they are not made in comparison with agreed standards. Generally, these should take the form of a hand-held key.
The measurement of yield is less controversial and is usually scored in terms of weight or number (117). However, the relation of disease severity to yield may not be obvious and requires empirical measurement. Various models—critical-point, multiple-point, and Area Under the Disease Progress Curve (AUDPC)—have been described (117). However, yield may be more directly related to the duration and amount of tissue that remains healthy (10). Integrated estimates of the relative damage done by diseases in different regions and environments, to arrive at comparative values for economic impact, are especially challenging (reviewed in 41, 92, 97, 106).
Identifying Sources of Inoculum and Its Means of Dissemination
Once the causal agent of a disease has been identified and its potential effects ascertained, either by reference to the literature or by experimentation, to be sufficiently significant to warrant control, the identification of sources of inoculum and its dissemination become immediate priorities. Infected seed, propagative material, or intact plants are often the means by which a pathogen may traverse long distances and establish itself in a new country. For example, race 3 biovar 2 of Ralstonia solanacearum was imported into the United States in infected geranium plants from Kenya and Guatemala, as noted above (http://www.aphis.usda.gov/ppq/ep/ralstonia/background.html). Similarly, blight of chickpea caused by Ascochyta rabiei was imported into the U.S. Pacific states and Australia on infected seed (40, 71, 76). Unfortunately, deliberate release of plant pathogens by terrorists cannot be ruled out (93).
Virulence Characteristics of Pathogens, Their Nature, Dissemination, and Evolution
Plant pathogens are in a constant state of flux. Not only do they move in space and time, but they also evolve. In particular, they evolve variants capable of overcoming the resistance of genotypes of crop plants that are widely grown and that may have been the results of years of work by plant breeders. It is essential, therefore, if we are to control these pathogens, that we understand the nature of pathogenicity, how it may be transferred between organisms, and how it may evolve.
For an organism to be a plant pathogen, it has to have a number of virulence functions such as the production of degradative enzymes (53, 59, 104, 136), toxins (9, 77, 134), and "effector" molecules (1, 78, 131). The last are the products of avr/pth genes. These are genes that are necessary for pathogenicity in host plants but confer avirulence in resistant plants. For example, in Xanthomonas citri the pthA gene was originally identified as a pathogenicity factor because it controls the ability of the bacterium to multiply in citrus plants and cause cankers. However, the same gene in X. campestris pvs. phaseoli and malvacearum confers avirulence on bean and cotton, respectively (84).
Interference with induced resistance is the primary target of some virulence factors. For example, a number of pathogens can convert the phytoalexins produced by their hosts to less toxic compounds. Genetic studies of populations of the pea pathogen Nectria haematococca have shown that metabolism of the phytoalexin pisatin is an important virulence mechanism of the fungus and that the genes required to cause disease in pea are clustered on a chromosome that is dispensable for normal growth (23).
Interference with the induction of defense may be an important virulence mechanism of bacterial pathogens (25). The hopPtoM and avrE genes of Pseudomonas syringae encode suppressors of salicylic acid-dependent callose deposition and promote cell death. Moreover, these genes are widespread in bacterial pathogens of plants, suggesting that they are important general virulence components (25). The attack on the signaling pathways of plant defense is likely to prove a fertile field of discovery in the future for those interested in virulence mechanisms of plant pathogens. For example, an isolate of Colletotrichum gloeosporioides metabolized salicylic acid in culture (2). Since this compound is fundamental to both genetic and systemic acquired resistance (26, 39), its destruction may go some way to explaining the wide host range of some strains of this pathogen.
Other genes necessary for pathogenicity or virulence have been uncovered by gene disruption experiments. For example, when genes encoding two proteins, ECP1 and ECP2, that are expressed during the infection of tomato by Cladosporium fulvum were disrupted, virulence was attenuated (81). ECP2 was the more important of the two since strains deficient in the production of this protein colonized tomato leaf tissue poorly and produced little emerging mycelium and few conidia.
Pathogenicity genes may be disseminated among potential plant pathogens by horizontal gene transfer. In bacterial plant pathogens, virulence genes and regulatory genes required for pathogenicity are often found in blocks, called pathogenicity islands, on the bacterial chromosome (78). These differ in G + C content from that of the bacterial species containing them, implying their acquisition from another organism. For example, the horizontal transfer of a pathogenicity island carrying the nec1 virulence gene and flanking sequences is thought to have occurred among Streptomyces strains in the Diastatochromogenes cluster (18). Moreover, it was postulated that the peptide synthase gene txtAB, which is required for the synthesis of a toxin, thaxtomin A, is located on the pathogenicity island and that this has conferred plant pathogenicity on saprophytic strains (18).
Horizontal gene transfer is also thought to occur in fungi (103) and may be an important mechanism for the acquisition of pathogenicity. For example, the pathogenicity cluster of N. haematococca (see above) is thought to have been acquired by horizontal gene transfer because its genes differ in G + C content and codon usage from genes in other chromosomes. Also, homologs have been found in another pea pathogen, Fusarium oxysporum f.sp. pisi, but are uncommon among other fungi that are closely related phylogenetically. Moreover, homologs have also been found in Neocosmospora boniensis, which is also able to cause disease in pea (124).
Genes required for the synthesis of host-selective toxins of strains of Alternaria alternata provide another example of possible horizontal gene transfer in fungi. Two genes, ATK1 and ATK2, from the Japanese pear pathotype of A. alternata were necessary for both production of the host-selective AK-toxin and pathogenicity for certain genotypes of pear (122). Homologs of these genes were found in strawberry and tangerine pathotypes of A. alternata, which produce toxins that are similar in structure to AK-toxin, but not in other pathotypes nor in nonpathogenic strains. AKT homologs have been found in multiple copies on a 1.05-Mb chromosome of a strawberry pathotype (51). The chromosome, like the one from N. haematococca that is responsible for the metabolism of pisatin (23), appeared to be conditionally dispensable since it was not essential for saprophytic growth.
So far virulence attributes of pathogens that are positive have been discussed. However, when considering gene-for-gene interactions, the absence or alteration of avirulence genes are important causes of virulence. For example, those races of the tomato pathogen Cladosporium fulvum that are virulent for tomatoes with the Cf 9 gene lack the corresponding avirulence gene avr9 (68). In contrast, three races of the pathogen that were virulent for tomatoes with the Cf 4 gene had Avr4 genes that were modified by a single base change, A to G, which occurred at a different position in each race. The result of the changes was that the proteins contained a tyrosine residue (codon TAT) rather than a cysteine residue (codon TGT). The modified proteins were unstable in the plant and therefore failed to trigger a resistance response (68).
Although loss or point mutations could explain the evolution of races of C. fulvum that are virulent to tomatoes with the Cf 9 and the Cf4 genes, respectively, other causes of mutation may also be at work with other plant-pathogen systems. The avrBs1 gene of the bell pepper pathogen X. campestris pv. vesicatoria, which causes bacterial spot of the plant, is one example. Here the gene is frequently disrupted by an insertion sequence, IS476, rendering the corresponding resistance gene, Bs1, ineffective. Ironically, selection for the insertion sequence was promoted by the use of copper sprays that were used to protect the plant against the disease, the reason being that they selected for bacteria containing a plasmid with a copper resistance gene that also contained three copies of the insertion sequence (74).
No doubt other virulence factors will be discovered in the future. Exactly what these are will become plainer in the next few years with the increasing use of microarrays to determine which genes are expressed during pathogenesis and gene disruption experiments (79, 89, 101, 114).
HOW CAN THE THREATS BE MINIMIZED?
Exclusion, Elimination, or Reduction of Inoculum
The most fundamental approach to the management of a disease is to ensure that it is not present through exclusion (quarantine) or eradication (28). National Plant Protection Services assume responsibility for protecting their countries from the unwanted entry of new pathogens and pests and for coordinating programs to eradicate those that have recently arrived and are still sufficiently confined for their elimination to be realistic. Regional plant protection organizations, for example, the European and Mediterranean Plant Protection Organization (EPPO), play a role in coordinating such "phytosanitary" activities across a region (115), and the Secretariat of the International Plant Protection Convention has a global responsibility in this field. While developed countries are sometimes concerned with the rare event of malicious introduction of pathogens as a form of bioterrorism, there is enormous scope for simply strengthening the resources and capacity of plant protection services in developing countries, with the goal of improving food security (3).
Plant material may be tested for specific pathogens, and a number of rapid and sensitive techniques are available (reviewed in 107, 109). Material that is found to be infected can then be destroyed. Once a pathogen is endemic in a region, much can be done to limit the amount of inoculum produced and its effectiveness. Seed and propagative material may be treated with biocides, and infected plants in the field may be rogued or the infected parts pruned (36, 128).
There are many other ways in which the farmer can take action to reduce disease in the field. Inoculum from crop residues may be reduced by crop rotation (in practice, this is one of the most important approaches to disease control), by plowing (118) or by adjusting planting time, and from the soil by solarization (73), the addition of amendments (83), and in some instances, by flooding (127). Some pathogens require specific wavelengths of light in order to sporulate (102). Their prevalence in greenhouses may be reduced by coating the glass with materials that do not allow the penetration of such wavelengths while still allowing through the light of wavelengths important for photosynthesis (102).
In the field, however, when an attack is imminent, there is little alternative to the application of pesticides, although by forecasting the occurrence of conditions favorable to the pathogen, the number of applications and the amount of pesticide used may be reduced (55). Many crops do benefit from routine or managed application of pesticides and this remains one of the principal control methods available for pathogens, especially fungi and nematodes, and their vectors, especially insects and nematodes (53a). In developing countries the use of pesticides is critically important for seed health. Pesticide use is particularly appropriate in systems of integrated pest management (94a), in which the aim is to create a sustainable balance of complementary approaches to disease control, avoiding excessive use of pesticides whose side-effects may be unwanted.
Some control of windborne pathogens may be achieved by planting mixtures of varieties or multilines. For example, an experiment on the control of rice blast caused by Magnaporthe grisea was carried out over a large area in Yunnan Province, China. Susceptible sticky rice was planted as single rows interspersed between four or sometimes six rows of resistant hybrid rice. Grain production of sticky rice was 89% greater and blast 94% less when planted in mixtures compared with monocultures (140). This principle may be extended to intercropping whereby different species of plants are grown in close proximity. However, care must be taken to ensure that no two or more members of the mixture are susceptible to the same organism, a condition that is not easy to fulfil with pathogens that have a wide host range. Diseases caused by pathogens such as mycoplasmas, viruses, and viroids are transmitted by vectors. It follows that control of vector mobility, for example by means of reflective mulches or interference with their feeding habits on plants by the use of genotypes with sufficient concentrations of antifeedants, would reduce infection by these agents (43, 47).
While there are many examples of successful use of biological control agents to control insect pests of crops, there are far fewer cases of successful use of such methods for field management of plant diseases (18a). Nevertheless, benefit is probably often achieved from unrecognized effects of biological control as when, for example, incorporation of crop residues into the soil affects the soil microflora to the disadvantage of a soilborne pathogen.
Rendering Crop Plants Genetically Resistant to Disease
One of the most effective approaches to disease management is the use of genetically resistant plants. Although the genetic basis for resistance to a plant disease was first elucidated only in 1905 (13), selection for disease resistance has probably been practiced by farmers and later by plant breeders since the beginning of agriculture. Spontaneous variation in disease resistance can be selected, and genes for resistance can be introduced by hybridization.
Genetic resistance to disease, once secured, is a low-cost method of control. Like other approaches to disease management, it need not be complete to be useful. Rather, it is more important that it should have the characteristic of durability, a concept proposed by Johnson (66). Durable resistance, i.e., resistance that remains effective over many years, can only be recognized retrospectively. It may not be readily available since pathogens have a facility, as described above, to evolve virulent variants, a phenomenon that has been much studied in terms of mechanism, genetics, and epidemiology. However, knowledge of the biochemistry and molecular biology of disease has identified some instances where durability may be expected (see below). Furthermore, examples of resistance that have been shown by experience to be durable can be used in breeding programs with the expectation that their effect will again be durable.
Advances in genomics have demonstrated that a considerable proportion, 1%–2%, of a plant's genome is devoted to resistance genes or genes with similar properties that could conceivably confer resistance to a pathogen possessing a complementary avirulence gene. Such genes are often clustered or occur in tandem repeats, suggesting that resistance genes with different specificities arise by gene duplication followed by intragenic and intergenic recombination, gene conversion, and diversifying selection (96). Maintenance of this genetic flux is crucial to the survival of the plant and therefore for maintenance in the gene pool from which the plant breeder can draw.
In gene-for-gene systems, resistance is triggered by the direct or indirect recognition of an avirulence gene product of the pathogen by a resistance gene product of the plant (16, 132). Breeding plants with resistance against a specific disease in such interactions requires the identification of resistant plants, which are then crossed with agronomically acceptable but susceptible plants. A program of backcrossing to the susceptible parent and selection of resistant phenotypes leads to the production of plants that are similar to the susceptible parent but have the required resistance. Typically, this process takes 10 or more years, and by this time, in some instances, the pathogen has already evolved a variant that is not recognized by the hybrid, leading to susceptibility.
Johnson (67) has described the challenge of breeding for resistance to two contrasting diseases of wheat: yellow (stripe) rust (caused by Puccinia striiformis) and eyespot (caused by Pseudocercosporella herpotrichoides). P. striiformis occurs as many races, differing genetically in their virulence to wheat varieties with different resistance genes, in a gene-for-gene relationship. The development of a new race can suddenly negate years of effort by a plant breeder, as happened in the United Kingdom to the variety Joss Cambier in 1971, causing fears of a crisis like the one due to southern corn leaf blight in the United States at the same time (see below), even though its slow-rusting quality had raised hopes of durability. Resistance to P. herpotrichoides is normally durable and not race-specific, but only moderate in degree. Introduction of a higher degree of resistance required cytogenetic manipulation to allow recombination between wheat and a wild relative from North Africa, Aegilops ventricosa, which possessed resistance. A block of genes was introduced into wheat that was undefined (as is normal in plant breeding) but contained one or more genes for resistance. Coming from an alien plant species, this might now be regarded with suspicion by some as being transgenic, although the procedure then appeared benign and a valuable application of novel genetic technology.
Marker-assisted selection (MAS) can help the breeder since it not only reduces the reliance on laborious and fallible screening procedures but also allows the pyramiding of resistance genes (60). Moreover, MAS may be used in the selection of genotypes with quantitative resistance to pathogens (17). The essential feature of any marker is that it should be tightly linked to the gene specifying the required character—in this case, resistance. For example, two microsatellite markers were found to cosegregate with two resistance genes for rice blast (38). Similarly, MAS is now routine in bean breeding programs for resistance to several diseases such as bean rust, anthracnose, common bacterial blight, and white mold (75).
Maintaining the Genetic Diversity of Crop Plants
Why is genetic diversity desirable in crops? An example will show how uniformity can lead to genetic vulnerability.
An unexpected and devastating epidemic of southern corn leaf blight (caused by Cochliobolus heterostrophus) occurred on the maize crop in the United States in 1970, resulting in an estimated loss of 1 billion bushels of the crop with a value of $1 billion (123, 130); adjusted for inflation, this would have a value of more than $4 billion in 2005. Apart from its use as human food, maize was the principal component of livestock feed, and it was a major export commodity. The impact was such that the U.S. Department of Agriculture and indeed U.S. President Nixon were drawn into reacting to what was seen as a potential agricultural crisis. Farmers' organizations in the U.S. corn belt brought legal actions against companies that had supplied their seed, for reasons that will become clear.
The epidemic proved to be due to the rapid evolution of a new race of the pathogen, Race T. This race is specific for maize that has a particular cytoplasm containing the gene T-cms and renders such crops highly susceptible to attack. The so-called Texas cytoplasm (T-cytoplasm) was widespread in 1970 as it contained a valuable mitochondrial gene that conferred male sterility and was consequently used in the production of hybrid seed.
Maize seed is normally produced as an F1 hybrid between different parents, because the hybrid vigor released in the F1 generation results in significantly enhanced performance. Maize is naturally self-pollinating, so some intervention is needed to produce F1 hybrid seed: for example, the pollen-producing tassels can be removed by hand. The male-sterilizing effect of T-cytoplasm was universally attractive as a means of reducing the cost of hybrid seed production. The plants were self-sterile so they did not need to be manually detasseled. This cytoplasmically based technology was indeed a revolution in plant breeding. It suited the seed companies, and it suited the farmers. These were the factors that led to at least 80% of the US maize crop being sown with cultivars based on the Texas cytoplasm by 1970.
The critical characteristic of Race T of C. heterostrophus is that it produces a toxin specific for the mitochondria of the T-cytoplasm, causing necrotic lesions on the leaves, stalks, and ears, which in turn produce inoculum that may lead to rapid spread of the disease. Symptoms can be severe, and as a result the plant may die and produce no useful yield.
The nearly universal use of cultivars with T-cytoplasm by 1970 selected strongly for Race T in the population of C. heterostrophus. It is unusual for cytoplasmic genes to affect resistance to disease, and it would not easily have been predicted that the widespread adoption of a particular cytoplasm would result in a new and damaging epidemic.
The scientific and public impact of the Race T episode provoked widespread concern about what was called genetic vulnerability (56). This valuable concept is meant to carry a warning that genetic uniformity leads to the potential for problems, if they occur, to be widespread and intense. Granted that it was difficult to predict the emergence of Race T, we might now think that it should have been easier to be forewarned that the very widespread use of T-cytoplasm rendered the maize crop vulnerable to something.
The term genetic vulnerability is less familiar now, and some related concepts that currently command greater attention, such as biodiversity and perhaps factory farming, do not carry the same warning that uniformity itself brings potential danger—especially from plant pathogens. It has been convincingly argued that genetic mixtures possess greater stability of performance and that their inherent resistance to disease is rendered more effective and more durable by their diversity (133). Yet uniform crops are convenient and financially efficient in the short term, and in practice they remain the norm. In most environments, a degree of genetic diversity is more readily achieved across the farm, or across a region, whether by accident or design.
Exploiting the Knowledge of the Biochemistry and Molecular Biology of Disease in Order to Increase Resistance
Plant pathogens are often at their most vulnerable at the pre-penetration stage of infection and at their most responsive to environmental influence. For example, Striga may be induced to germinate in the absence of the host by the introduction of ethylene into the soil, although this is a hazardous procedure. However, a strain of Pseudomonas syringae pv. glycinea has been found that produces ethylene and that consistently stimulated better germination of seed of the parasite than the gas (11). This discovery holds out the prospect that amendment of soil with this organism might lead to control of Striga owing to its ability to stimulate germination of seed of the parasite in the absence of a suitable host.
Other procedures for enhancing resistance include exploiting a pathogen's dependence on certain virulence mechanisms. For example, X. albilineans, causal agent of sugarcane leaf scald, produces a family of toxins and antibiotics known as the albicidins, which selectively block DNA replication in bacteria and chloroplasts. In consequence, sugarcane infected by the bacterium is chlorotic. During a screening program, an albicidin-resistant isolate of Pantoea dispersa was found that efficiently detoxified albicidin and was effective as a biocontrol agent (137).
As mentioned above, the secretion of two proteins, ECPI and ECP2, by Cladosporium fulvum was required for full expression of virulence of this tomato pathogen. When a construct of the gene encoding ECP2 was made with potato virus X (PVX::ECP2) and used to infect a range of tomato genotypes, four were found that responded with a systemic hypersensitive response. This reaction was encoded by a single dominant gene, designated Cf-ECP2, that recognizes the protein ECP2. Since this factor is required for virulence, the resistance conferred by Cf-ECP2 is expected to be durable (82).
The GM Option
With the recognition that tumor induction in plants by Agrobacterium tumefaciens is caused by the transformation of its hosts with T-DNA present on a plasmid in the bacterium, the use of this vector to transform plants has become almost routine, although other techniques such as electroporation and biolistics are also used. The question then arises as to what genes should be considered for introduction into crop plants. Given the severe limitations of agricultural technology, especially in developing countries, to provide secure food supplies for a rapidly growing global population, the opportunity to introduce entirely new characteristics into plants has been seized upon energetically by plant breeders, through transgenic techniques—often described as genetic modification (GM). Disease resistance is among the most promising applications of GM. However, the growth of transgenic technologies in agricultural practice has been limited by public opinion in some countries. Whatever precautions are taken in their development and testing, transgenic plants are regarded by some as unnatural and potentially dangerous. This climate of opinion, especially in developed countries, has unfortunately limited the adoption of novel and potentially valuable opportunities for crop improvement—especially in developing countries, where new developments are most needed to enhance food security.
This attitude could change as the benefits of transgenic technologies become more apparent in practice. Indeed, during the period 1996 to 2003, the global area of transgenic crops increased 40-fold, from 1.7 to 67.7 million ha. Furthermore, 30% of this area is grown in developing countries, where the recent rate of increase has been higher than in industrialized countries (64).
An interesting discussion of the potential of biotechnology for solving intractable constraints to food security in Africa is provided by Gressel et al. (48) and, in a more general approach, by Christou & Twyman (20). Candidate genes include those that inhibit or destroy virulence attributes of the pathogen, such as degradative enzymes and toxins, and those that enhance defense. The latter include genes that increase the concentrations of saponins, antimicrobial peptides, and reactive oxygen species or modify the phytoalexin response, and those that switch on systemic acquired resistance. Paradoxically, transformation of plants with pathogen genes such as coat protein and replicase genes of viruses confers resistance, a phenomenon known as pathogen-derived resistance. Finally, the ability to transform plants allows the pyramiding of resistance genes, including those that are normally found as alleles at the same locus (117). Space does not permit the extensive discussion of all these possibilities, so mention will only be made of some of the more recent and most promising new developments.
Apple has been transformed with a stilbene synthase gene from grapevine and a polygalacturonase-inhibiting protein (PGIP) from kiwi (120). The stilbene synthase catalyzes the synthesis of resveratrol, a phytoalexin found in both grapevine and peanuts but not apples. It is therefore probable that apple pathogens would be sensitive to it since they would be unlikely to have evolved a mechanism for detoxifying the compound. Chemical analysis demonstrated the presence of a resveratrol glycoside in transformed plants, and the presence of the PGIP gene was demonstrated by PCR and Southern blot analysis. Although studies of the disease resistance of the transgenic apples are still awaited, precedents in the literature suggest that they will be more resistant to fungal pathogens than the untransformed types (49, 100).
Transformation of rice with the afp gene of Aspergillus giganteus, which encodes the antifungal AFP protein, has been achieved and the gene demonstrated to be stably integrated and inherited. Protein extracts of the transgenic plants were antifungal, and detached leaves from the plants showed varying levels of resistance to rice blast caused by Magnaporthe grisea (21). In other experiments with rice, overexpression of a thionin gene from oats had resistance to Burkholderia plantarii that was virtually complete (62).
As already mentioned, an isolate of Pantoea dispersa has been found that was an effective biocontrol agent for sugarcane leaf scald caused by X. albilineans, probably owing to its ability to degrade the albicidin toxins produced by the pathogen (137). Sugarcane was transformed with the gene responsible for toxin degradation. Transgenics, expressing the gene at levels of 1–10 ng of enzyme per mg leaf protein, did not develop the chlorotic symptoms that are characteristic of the disease when challenged with the pathogen, whereas the controls were severely affected (138).
Peanut cultivars transformed by microprojectile bombardment of embryogenic callus derived from mature seeds with the coat protein gene of peanut stripe potyvirus gave rise to plants that were resistant or immune to the virus. The transgenics are now an important germplasm resource for countries where peanut stripe virus is endemic (54). Farming of papaya (Carica papaya) in Hawaii was threatened by papaya ringspot virus. Initially, control was obtained by cross protection with a mild strain of the virus (44). However, more recently a transgenic approach has been adopted in which plants have been transformed with the coat protein of the virus (6, 44). Although some coat protein genes tended to give high levels of protection only to homologous strains of the virus, others gave broad-spectrum resistance (6, 7, 37).
There is a vigorous global debate about the scientific and ethical acceptability of transgenic crops—those that are genetically modified through biotechnology. It will probably seem surprising in the future that this caused so great a cultural divide at the turn of the twenty-first century. But what is beyond doubt is that, in these early stages of a revolution in the technology of plant improvement, the number genes being deployed is small.
Transgenic crops in commercial production (currently maize, soybean, cotton, and canola) occupied 68 Mha in 18 countries in 2003, and this rose by 15% from 2002 (64). At present, only two traits are the subject of the commercialized transgenic biotechnology: herbicide (glyphosate) tolerance, and insect resistance conferred by the Bt gene from Bacillus thuringiensis. This means that new technologies to fight weeds and insects are in place, in itself a remarkable and positive step. Technologies have to start somewhere, and this has started with just two traits. The time will soon come when there are many genes for insect and disease resistance in play, and then we may feel more secure from the genetic vulnerability that may be presented at the current stage of development of the technology.
THE CHALLENGE OF THE FUTURE
The challenge of the future may be viewed at two levels, the biological and the political. At the biological level there is clearly much to be done to expand our knowledge of plant pathogens and our ability to deal with them. At the political level, plant pathologists should persistently draw these opportunities to the attention of politicians and public and private fund holders until they make appropriate provision for the subject, which is vital to the support of human life in industrialized and especially in developing countries.
The Biological Challenge
Plant pathogens pose an enormous problem for global food security. In the first instance we are still ignorant of the identity of many damaging pathogens that currently exist. Second, our means of measuring the severity of the symptoms they cause is often subjective and qualitative rather than objective and quantitative. Third, there are few examples in which a definite and dependable coefficient has been established that links the severity of symptoms to failure to reach obtainable yields. Fourth, plant pathogen populations are variable in time and space. Although there have been many epidemiological studies, much remains to be done in the field of limiting inoculum, its multiplication, effectiveness, and spread. Finally, and potentially most dangerously, plant pathogens are genetically variable. Nobody predicted the occurrence of the recombination of African Cassava mosaic virus (ACMV) with East African Cassava mosaic virus (EACMV) to produce a strain, UgV. Nor was it predicted that the UgV strain, in combination with ACMV in doubly infected plants, would give rise to an unusually severe form of the disease causing numerous farmers in Africa to abandon growing this vital staple crop. Also, nobody predicted that the near-exclusive reliance on the T-cms gene for male sterility in maize in the United States would presage the epidemic of southern corn leaf blight in 1970, one of the worst plant disease epidemics ever recorded. This stands as a warning that genetic uniformity of even a small part of the genome of a widely grown crop risks the hazard of a major epidemic of a plant pathogen with the ability to exploit the trait(s) that it governs.
It is difficult to predict the origin of the next plant disease catastrophe that will affect one or other of our crops vital to food security in some part of the globe. However, of two things we can be certain: There is inadequate control of known diseases in many parts of the world today, and in the future there will be some unpleasant shocks from pathogens that have evolved new virulence characteristics.
The Political Challenge
In many countries, politicians and their constituents are preoccupied by the threat to peace and prosperity posed by terrorism. It is time that they were also concerned with the terrorism perpetrated by plant pathogens and pests on our sources of food, which threaten our very security and survival.
There are no easy solutions to poverty. Scientists can, however, contribute in important ways to the more tangible issue of food security. For plant pathologists, the loss of food to plant disease presents a challenge. Experience shows that there is always some new threat to food security, so the challenge is ongoing. The people most strategically placed to meet that challenge on behalf of the undernourished are the staff of the national programs in the affected countries. But these are precisely the institutions that are under-resourced to meet the challenge.
This dilemma is somewhat relieved by the organizations with resources from the developed world that have a concern for food security in developing countries, and by their support for national programs there. The Consultative Group for International Agricultural Research is of primary importance, with its mission to bring the benefits of modern science to poor farmers "living on the fringes of the global economy" (22a). Eleven of its globally distributed International Agricultural Research Centers have programs in the management of plant disease, each Center being mandated to handle certain crops: Africa Rice Center (WARDA), Centro Internacional de Agricultura Tropical (CIAT), Center for International Forestry Research (CIFOR), Centro Internacional de Mejoramiento de Maiz y Trigo (CIMMYT), Centro Internacional de la Papa (CIP), International Center for Agricultural Research in the Dry Areas (ICARDA), International Crops Research Institute for the Semi-Arid Tropics (ICRISAT), International Institute of Tropical Agriculture (IITA), International Plant Genetic Resources Institute (IPGRI), International Rice Research Institute (IRRI), World Agroforestry Centre (ICRAF).
Internationally, the Food and Agriculture Organization (FAO) of the United Nations plays a key role in addressing the challenge of food security, including the depravations of plant disease (34). And throughout the developed world, national governments have development-assistance agencies that support agricultural development in developing with countries, sometimes with specific attention to plant disease issues.
At a voluntary level, the International Society for Plant Pathology (ISPP) is among many scientific organizations that are deeply concerned for global food security and the contribution that science can make to it. Being involved in ISPP's programs, we welcome participation in the Society's small effort to address these issues through a Task Force on Global Food Security (http://www.isppweb.org/foodsecurity_background.asp). We present this as a small example of an action-oriented project designed to address pressing local issues. The initiative was stimulated by a public meeting on "Global Food Security: the Role for Plant Pathology" during the seventh International Congress of Plant Pathology in Edinburgh in 1998. It was recognized then that more than 800 million people do not have adequate food, that 1.3 billion live on less than $1 a day, and that at least 10% of global food production is lost to plant disease. The Task Force has initiated activities in (a) changing public policy and opinions on global food security; (b) postgraduate training for plant pathologists in developing countries; (c) quantification of the economic impact of some major diseases; (d) farmer training in simple disease management: pilot project in Ghana; and (e) development of the ISPP website.
The project in Ghana is on development of appropriate strategies to control cassava diseases. Its aims are to (a) develop appropriate measures to control root rot disease of cassava; (b) create and increase farmers' awareness of diseases of cassava, particularly the root rot caused by Polyporus sulphureus; (c) train and equip farmers with simple skills that will enable them to identify diseases and control them on their farms; and (d) develop simple educational materials on diseases and their control to improve extension delivery. Those expected to benefit include over 80% of 18 million people in Ghana who depend on cassava as a major staple food, as well as industries that depend on cassava as a raw material.
The efforts of an international society with voluntary officers and limited funding can only be sadly inadequate in relation to the enormity of the problem of providing food for the world and securing its availability into an uncertain future.
The Annual Review of Phytopathology is online at http://phyto.annualreviews.org
Securing Food for the World: Vision and Perspectives
Readers may be interested in "Vision - World Nutrition 2020", taken from a presentation at EXPO 2000, Hannover, Germany, prepared by the Editor of ISPPNews, Prof. Dr. Joseph-Alexander Verreet, Christian-Albrechts-University of Kiel, Institute of Phytopathology, Hermann-Rodewald-Str. 9, D-24118 Kiel, Germany. Click here to download .pdf file (1.5 MB). Comments are welcomed by the Editor. E-mail: javerreet@phytomed.uni-kiel.de.
ACTIVITY 2: ENHANCED PhD TRAINING FOR PLANT PATHOLOGY IN DEVELOPING COUNTRIES
Mike Jeger had no results to report.
Richard Strange was thanked for his willingness to undertake a questionnaire survey on the provision of Ph.D. and M.Sc. courses in plant pathology for students normally domiciled in developing countries.
This was conducted after the Congress and an analysis of the results is available under the title "Provision of Ph.D. and M.Sc. Courses in Plant Pathology for Students Normally Domiciled in Developing Countries".
ACTIVITY 3: QUANTIFICATION OF THE ECONOMIC IMPACT OF SOME MAJOR DISEASES
Jill Lenne could not be present to report. She is expected to prepare a paper on the subject.
ACTIVITY 4: FARMER TRAINING IN SIMPLE DISEASE MANAGEMENT: PILOT PROJECT FOR CASSAVA IN GHANA
Emmanuel Moses could not be present to report. His two papers were examined on:
• Report for ISPP Task Force on Global Food Security. Activity 4: Farmer Training in Simple Disease Mangement on Cassava in Ghana.
• Factsheet: Control of Cassava Diseases.
His achievement in delivering these reports was appreciated by the Task Force.
ACTIVITY 5: DEVELOPMENT OF THE ISPP WEBSITE
Peter Scott reported, with a demonstration of some key pages from ISPP's new Website, redesigned for the Congress at
www.ISPPWeb.org
3. CONGRESS CHALLENGE
The Task Force evaluated four Proposals received for the Congress Challenge, listed below.
3.1. Mark Holderness January 10, 2003
Improving seed health and quality in staple crops for the resource-poor
3.2. Chrys Akem January 17, 2003
Empowering Farmers to Manage Crop diseases for Increased Food Security in Developing Countries
[3.3. Claude Fauquet January 17, 2003. A commentary, not a project proposal]
Use ISPP "political" power, and its resources, to enhance research funding in this field of science.
3.4. Hubert Zandstra January 20, 2003
Reducing Risk of Potato Production in South West Uganda
3.5. Emmanuel Moses January 20, 2003
Development of Appropriate Strategies to Control Cassava Diseases in Ghana
After much consideration, the Task Force recommended the Proposal by Emmanuel Moses on "Development of Appropriate Strategies to Control Cassava Diseases in Ghana" for support under the Congress Challenge initiative. This was to be proposed for endorsement by the Executive Committee of ISPP, and was subject to resolution of some issues identified during consideration of the Proposal by the Task Force.
4. NEW INITIATIVES
4.1. Review of Task Force activities, and general discussion
The paper by Claude Fauquet (3.3 above) was examined.
The view was noted and appreciated that ISPP's main role in the context of global food security should be to lobby for increased research funding.
4.2. Re-Focusing the Priorities
The paper from Chrys Akem was examined.
The views expressed were appreciated, including the endorsement of the continued relevance of the Task Force. The benefits of moving to a regional focus were questioned, but the gains to be achieved by enhancing interaction between North and South, for example by improved integration of PhD training, were endorsed. The benefit of improved collaboration between national societies was also supported.
5. ANY OTHER BUSINESS
Peter Scott agreed to remain as Chairman of the Task Force for the time being.
Richard Strange agreed to take over Activity 2 from Mike Jeger.
A report on Task Force activities, including recommendations of this meeting, would be posted on ISPPWeb and disseminated through ISPP-LIST.
6. NEXT MEETING
The Task Force should be active by e-mail correspondence. It should aim to meet at least once before the ICPP 2008, and then again at ICPP 2008.
This record of the meeting compiled by Chrys Akem, edited by Peter Scott

;){kind=link}